8月18日,国际学术期刊Zoological Research在线发表了中国科学院分子细胞科学卓越创新中心陈洛南组与西北工业大学王文组最新研究成果“Coevolutionary insights between promoters and transcription factors in the plant and animal kingdoms”。通过对跨越十亿年进化史,总共数百个动植物物种的启动子序列和转录因子理化性质分析,发现启动子和转录因子在动植物细胞中存在不同的共演化模式。该成果被选为当期的封面文章。
动物和植物从原始的单细胞生物(绿色鞭毛生物)分化(diverge)并各自持续地演化(evolve),促成了地球上自然生态的多样性(diversity)。在真核生物(eukaryotes)的基因转录(transcription)和调控(regulation)过程中,启动子(promoters)和转录因子(transcription factors)以及它们的相互作用扮演着决定性作用。然而,启动子和转录因子在物种水平上的演化轨迹及演化模式目前尚不清楚。
通过对跨越十亿年进化史,总计420个动物物种、223个植物物种的基因组序列分析和蛋白组理化性质分析表明:(1)序列的GC含量(GC-content)和蛋白质的等电点(isoelectric points)可分别作为基因的启动子和转录因子的演化特征印记(signatures)(Fig.1);(2)在动物界,启动子的GC含量和转录因子的等电点,在漫长的演化历程中,表现出持续的同步增长(Fig.2A,C);(3)在植物界,启动子的GC含量和转录因子的等电点,则表现出同步的降低趋势(Fig.3A,B)。启动子和转录因子的演化轨迹为“哺乳动物和鸟类一样,都是从爬行动物直接演化而来”提供了新的证据(Fig.2A,C,D,E,F)。
进一步通过分析启动子和转录因子之间相互作用的模式,以及它们之间演化轨迹的强相关性和显著性,表明启动子和转录因子在动物界呈现拮抗的共演化(antagonistic coevolution)(Fig.4A,B),而在植物界则呈现互利的共演化(mutualistic coevolution)(Fig.4C,D)。因为拮抗的共演化在所有共演化模式中更具有群体自然选择优势,这或许能解释,在现有的自然界中动物物种的多样性远高于植物物种(动物物种数大约有 777万,而植物只有29.8万,前者是后者的26倍)。该研究为探究启动子与转录因子间的相互作用机理,进一步理解动植物的分化、趋异演化及生态多样性提供了新的洞见(Fig.5)。
中国科学院分子细胞科学卓越创新中心陈洛南研究员、西北工业大学王文教授和中国科学院分子细胞科学卓越创新中心张劲松副研究员为该文的共同通讯作者。中国科学院分子细胞科学卓越创新中心张劲松副研究员、夏洁助理研究员和上海交通大学附属医学院王海权博士为共同第一作者。这一工作得到科技部重点研发计划,中科院战略性先导科技专项,国家自然科学基金等项目的支持。
文章链接:https://doi.org/10.24272/j.issn.2095-8137.2022.111

封面图片:Promoters and transcription factors enjoy antagonistic coevolution in the animal kingdom, but mutualistic coevolution in the plant kingdom.
封面设计:Jingsong Zhang, Libin Wu, LetPub
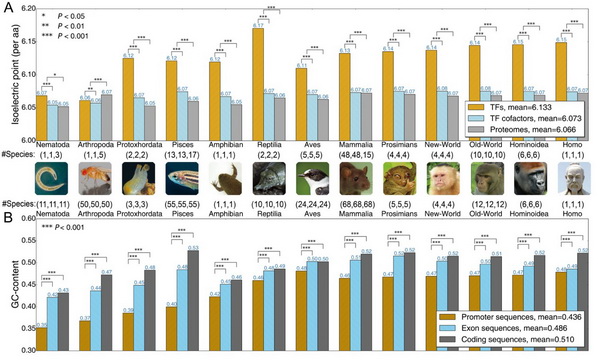
Fig.1 启动子与转录因子特征印记(signatures)。A:通过物种的转录因子、转录辅助因子、蛋白组序列的等电点(带电特征)分析,发现(1)转录因子与转录辅助因子之间具有显著差异;(2)转录因子与转录辅助因子作为整体与蛋白组之间具有显著差异。B:通过物种的启动子区、外显子区、编码区的GC-content分析,发现(1)启动子与外显子和编码区均具有显著差异;(2)启动子区域的具有更小的GC-content。因此,GC-content和等电点可分别作为启动子和转录因子的特征印记。
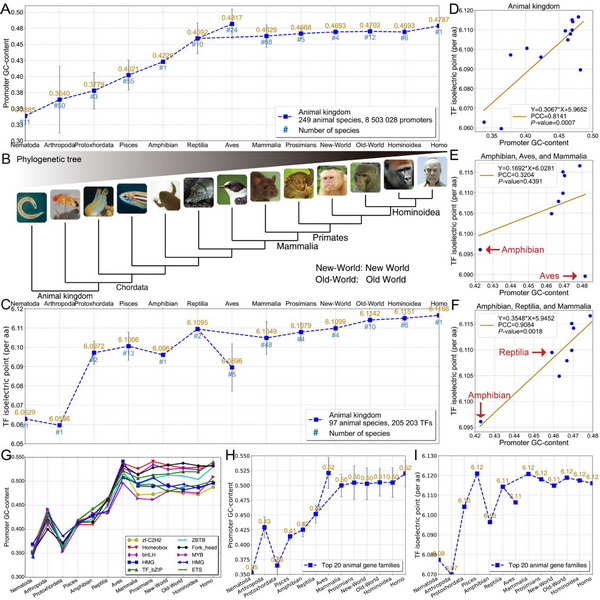
Fig.2 在动物界中,启动子和转录因子的共演化轨迹。A:从线虫到现代人249个物种的基因组的启动子区域的GC-content分析,发现随着物种演化从简单到复杂方向,GC-content呈现平滑的上升趋势(鸟类除外)。C:从线虫到现代人97个物种的蛋白组的转录因子(含辅助因子)区域的带电特征分析,发现随着物种演化从简单到复杂方向,等电点呈现上升趋势(鸟类除外)。D:通过启动子GC-content与转录因子等电点的相关性分析,发现两者具有强相关性和显著性。E:将两栖类、鸟类和哺乳类为一组,对GC-content和等电点进行相关性分析,发现这三个集合不具有相关性。F:将两栖类、爬行动物和哺乳类为一组,发现这三个集合具有强相关性和显著性。通过D、E、F分析,识别出鸟类是离群点(outlier),由此,为“哺乳动物是从爬行动物而不是从鸟类演化而来”提供了新的证据。G、H、I:在基因家族的启动子GC-content分析,发现与A图具有相似的演化轨迹。综上,在动物界中,启动子GC-content和转录因子等电点同步上升,具有演化上的强相关性和显著性。
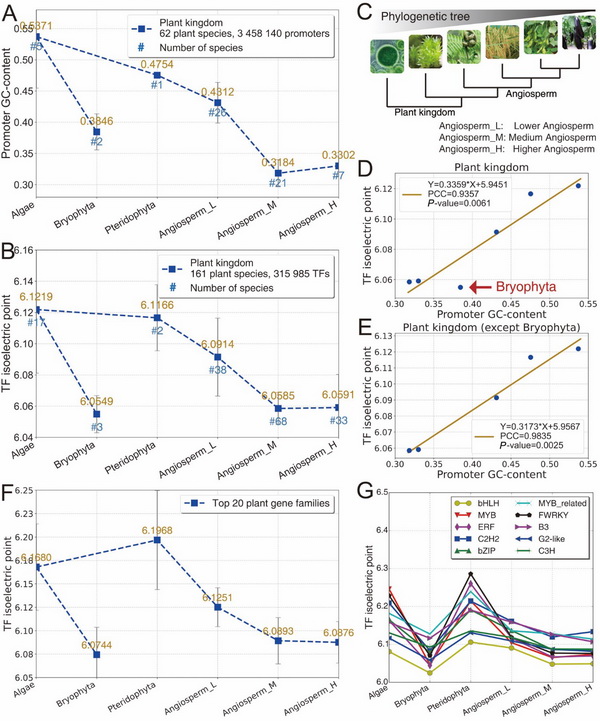
Fig.3在植物界中,启动子和转录因子的共演化轨迹。A:从藻类到被子植物,启动子的GC-content呈现下降趋势。B:从藻类到被子植物,转录因子的等电点也呈现下降趋势。D:从藻类到被子植物,启动子GC-content与转录因子等电点具有强相关性和显著性。E:将苔藓植物物种除外,从藻类到被子植物的演化过程中,启动子的GC-content与转录因子的等电点具有更强的相关性和显著性,由此为“苔藓植物是蕨类植物退化产生”学说提供潜在的证据。F、G:在基因家族层面,启动子GC-content具有与A图相似的演化轨迹。综上,在植物界中,启动子GC-content和转录因子等电点同步下降,具有演化上的强相关性和显著性。
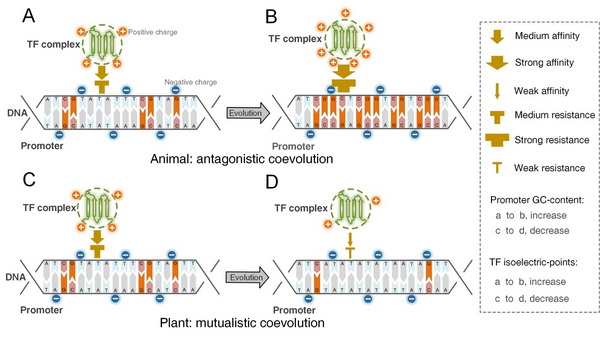
Fig.4 启动子和转录因子在动植物界中不同的共演化模式。在DNA双螺旋中,G/C之间具有三条化学键,A/T之间具有两条化学键,因此,启动子的GC-content影响其稳定性,GC含量越高双螺旋越稳定。另外,GC含量越高的区域,通常甲基化率越高。DNA双螺旋骨架的外侧通常带负电,表明转录因子带正电荷越多,越容易靠近双螺旋,进而与DNA发生相互作用。A 到 B:动物界中从简单到复杂的演化方向上,启动子GC-content递增,转录因子带正电荷量也同步递增。在演化进程上,启动子因GC-content的增加而更加稳定,转录因子若要靠近DNA双螺旋并从启动子区域的转录起始位点打开,则自身必须带有更多的正电荷。转录因子只有打开启动子并进行后续的转录和翻译,才会有转录因子“子代”产生。启动子一方面要保持自身的稳定性,又要产生新的“子代”,则必须依赖转录因子的调控。转录因子(寄生)对启动子(宿主)的寄生,并且互利关系,表明在动物细胞中,启动子和转录因子之间形成拮抗的共演化模式。C 到 D:植物界中从简单到复杂的演化方向上,启动子GC-content递减,转录因子带正电荷量也同步减少。在演化进程上,启动子因GC-content的降低而更容易被转录因子打开,当转录因子不需要更多的正电荷时,它们便呈现正电荷带电量减小趋势,两者利他的相互作用,表明植物细胞中,启动子和转录因子之间形成互利的共演化模式。 总之,启动子和转录因子在动物界呈现拮抗的共演化,而在植物界则呈现互利的共演化。
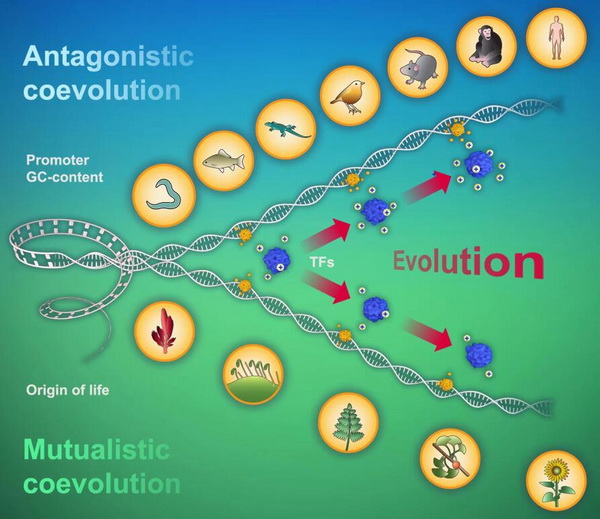
Fig.5 动植物界中启动子和转录因子共演化封面。图上方DNA双螺旋伸展趋势代表动物演化过程中启动子GC-content上升趋势;图下方DNA双螺旋伸展趋势代表植物演化过程中启动子GC-content下降趋势。蓝色的团状代表转录因子,转录因子周围的“+”号代表带正电量,伴随着演化进程,动物细胞中转录因子正电荷在递增,而植物细胞中转录因子正电荷在递减。动植物中启动子与转录因子不同的演化趋势,表明拮抗和互利两种不同的共演化模式。在原始的单细胞生物中,启动子和转录因子之间不同的演化模式,或是原始的单细胞生物分化为植物界、动物界等界的驱动力。拮抗的共演化模式具有群体自然选择的优势,因此,动物细胞中启动子和转录因子拮抗的共演化,或是驱动动物物种比植物物种具有更高的物种多样性。