北京时间4月4日,国际学术期刊Cell在线发表了中国科学院分子细胞科学卓越创新中心(生物化学与细胞生物学研究所)周斌研究组的最新成果“Tracing the origin of alveolar stem cells in lung repair and regeneration”。该研究建立了追踪肺上皮细胞的Cre-loxP和Dre-rox双同源重组酶介导的遗传谱系示踪新技术,结合多种小鼠肺脏损伤模型,系统研究了肺泡上皮干细胞的再生起源并揭示其细胞命运调控的分子机制。该工作不仅为肺脏疾病的临床治疗研究提供了新的科学依据和突破口,也为器官发育和再生医学研究提供了新技术和新方法。
肺作为一个复杂的多功能器官,对人类生存至关重要。肺上皮干细胞在肺脏再生修复中发挥重要作用,是肺脏再生和肺疾病领域的研究热点。以小鼠肺脏为例,肺叶内部主要包括支气管和肺泡两个区域。不同区域的肺上皮细胞种类和数目不同,并且含有各自的干细胞负责本区域上皮的稳态维持及损伤修复。支气管上皮主要由棒状细胞(Club cell,分子标记为Scgb1a1)、纤毛细胞(Ciliated cell)和神经内分泌细胞(Neuroendocrine cell, NE cell)等细胞构成,其中Club细胞和NE细胞属于干细胞,能维持支气管上皮更新;肺泡上皮主要由I型肺泡上皮细胞(Alveolar type I cell, AT1 cell)和II型肺泡上皮细胞(Alveolar type II cell, AT2 cell,分子标记为Sftpc)构成,其中AT2细胞作为肺泡干细胞能自我更新并分化为AT1细胞;在支气管和肺泡区域交界的位置,还存在一群支气管肺泡干细胞(Bronchioalveolar stem cells, BASCs),具有同时表达club细胞分子标记Scgb1a1和AT2细胞分子标记Sftpc的特点,周斌团队前期研究揭示了BASCs在肺损伤后具有向支气管和肺泡上皮双向分化的潜能。
AT2细胞作为肺泡上皮干细胞在肺泡损伤修复中发挥重要作用。因此鉴定和阐明AT2肺泡干细胞的再生起源及分子调控机制对肺脏疾病的治疗具有重要意义。近年来陆续有重要研究成果报道,发现在肺脏受损后,AT2细胞除了来源于自我更新外,还会起源于AT1细胞和club细胞,这些关于肺脏干细胞突破性的研究发现,极大推进了肺脏再生领域的发展。然而由于肺干细胞研究领域内广泛使用的传统谱系示踪技术存在非特异性标记的缺陷,导致这些重大发现一直存在科学争议,因此亟需构建谱系示踪新技术对这些科学争议一一进行阐明。周斌团队长期致力于小鼠体内谱系示踪新技术的开发和应用,曾利用双重组酶介导的谱系示踪技术解决了多个领域内重大科学问题,有力地推动了相关研究领域的发展和进步。
为了阐明AT2细胞的再生来源,研究人员首先对传统的单同源重组酶驱动的谱系示踪工具进行了系统性鉴定。Hopx-CreER是肺干细胞领域内普遍使用的AT1细胞标记示踪工具,对成体Hopx-CreER;R26-tdT工具小鼠进行肺切除手术(Pneumonectomy,PNX)或博来霉素(Bleomycin)诱导肺损伤后,发现标记的Hopx+细胞确实可以转分化为AT2细胞。然而对稳态下Hopx-CreER;R26-tdT工具小鼠的肺脏切片进行免疫荧光染色及统计分析后,发现此遗传工具除了高效率标记AT1细胞外,还“异位”或者“非特异”标记~66% club细胞、~12% BASCs、~92% ciliated细胞,还有极少量AT2细胞(~0.24%)。由于BASCs、club细胞和AT2细胞本身已被报道可以作为干细胞在肺损后贡献AT2细胞,因此会对Hopx-CreER;R26-tdT的谱系示踪结果产生很大干扰。为了探究Hopx-CreER的非特异性标记是否由于Hopx一条等位基因敲除引起的,研究人员构建了Hopx-2A-DreER。通过对Hopx-2A-DreER;R26-RSR-tdT工具小鼠肺脏进行检测,同样的发现tdTomato除了标记AT1细胞外,还“非特异”标记ciliated细胞、club细胞和少部分BASCs。研究人员进一步对另一种领域内使用的AT1遗传工具Ager-CreER进行鉴定,发现该工具除了高效率标记AT1细胞外还会“异位”标记一部分AT2细胞和少量BASCs。通过以上实验,研究人员揭示基于Hopx基因或Ager基因的传统单同源重组酶驱动的谱系示踪技术具有非特异性标记问题,因此无法利用其对AT1细胞进行严格的谱系示踪研究。
得到以上结果后,研究人员意识到领域内迫切需要构建谱系示踪新技术来实现对AT1细胞的特异性标记,于是提出利用双同源重组酶驱动的谱系示踪新技术来解决此难题。通过对比,研究人员发现Hopx-2A-DreER与Ager-CreER非特异性标记的细胞类型具有“互斥”特点,即Hopx-2A-DreER除了标记AT1细胞外会“异位”标记ciliated细胞、club细胞和少量BASCs,而Ager-CreER除了标记AT1细胞外会“异位”标记AT2细胞和少量BASCs。基于这个“互斥”特点,研究人员首先通过交叉标记策略(Intersectional strategy)构建了双同源重组遗传工具:Hopx-2A-DreER;Ager-CreER;R26-RSR-LSL-tdT,只有同时发生Dre-rox和Cre-loxP重组的细胞中,报告基因tdTomato才会表达,进而实现对Hopx+Ager+ AT1细胞特异性标记。结合多种肺损伤模型(肺切、博来霉素和高氧损伤),研究人员发现在肺损后AT1细胞不会转分化为AT2细胞,AT1细胞属于终末分化细胞。研究人员进一步通过嵌套策略(Nested strategy)构建了另一种双同源重组遗传工具:Hopx-2A-DreER;Sox2-CreER;Sftpc-CreER;R26-NR2。在此策略中,Hopx+AT1细胞会被特异性标记为ZsGreen,而“非特异”标记的club细胞、ciliated细胞、BASCs及AT2细胞均会被统一标记为tdTomato。然而结合多种肺损伤模型,研究人员仍然未检测到标记的AT1细胞转分化为AT2细胞。通过以上两种双同源重组谱系示踪新技术,研究人员揭示在肺损后AT1细胞不具备可塑性,不会转分化为AT2细胞。
研究人员进一步指出领域内club细胞遗传工具Scgb1a1-CreER同样具有非特异性标记问题,Scgb1a1-CreER;R26-tdT除了标记club细胞外还会“异位”标记BASCs和少量AT2细胞,这会对club细胞的谱系示踪结果产生干扰。因此研究人员基于Cre-loxP和Dre-rox构建了另一种双同源重组谱系示踪新策略,即Scgb1a1-CreER;Sftpc-DreER;R26-TLR遗传工具,解决以上非特异性标记问题。在此系统中club细胞内会发生Cre-loxP重组被标记为tdTomato;AT2细胞内会发生Dre-rox重组被标记为ZsGreen;BASCs内同时发生Cre-loxP和Dre-rox重组会被标记为tdTomato和ZsGreen,从而展现出黄色荧光标记。因此,该新技术可以实现对club细胞、BASCs和AT2细胞这三种细胞群同时特异性遗传标记,分别称之为Club-tracer、AT2-tracer和BASC-tracer。结合博来霉素损伤模型,研究人员发现肺损后club细胞和BASCs会大量分化为肺泡上皮(包括AT2和AT1细胞)进而促进肺泡再生,对肺泡损伤区域进行统计,显示~27%和~11%新生AT2细胞分别起源于club细胞和BASCs。为进一步挖掘club细胞对肺泡上皮修复的潜力,研究人员计划通过诱导BASCs和AT2细胞过表达p21蛋白封闭细胞增殖能力,使之不能参与再生修复,以此激发club细胞的可塑性潜力。为了实现这一目的,研究人员设计了一种新的双同源重组系统,即Sftpc-DreER;R26-rox-p21-GFP;Scgb1a1-CreER;R26-LZL-tdT遗传工具,club细胞内发生Cre-loxP重组被标记为tdTomato;AT2细胞内会发生Dre-rox重组被标记为GFP并表达p21蛋白;BASCs内同时发生Cre-loxP和Dre-rox重组会被标记为tdTomato和GFP并表达p21蛋白。对此系统成功验证后,研究人员随即结合博来霉素损伤模型来探究club细胞的可塑性。收取损伤4周后的肺脏进行分析,发现AT2细胞和BASCs原本的修复能力成功被抑制,并发现club细胞大量转分化为新生AT2细胞和AT1细胞,统计显示损伤区域~47%AT2细胞起源于club细胞。研究人员进一步分析了长时程(损伤后8个月)修复的肺脏,惊奇的发现与4周相比,club细胞贡献的E-cad+肺泡上皮(包括AT2细胞和AT1细胞)比例显著性上升,统计显示~82% AT2细胞起源于club细胞。在损伤严重的肺叶中,club起源的肺泡上皮细胞可以覆盖绝大部分肺泡区域。以上结果说明肺损后当AT2细胞和BASCs修复能力被抑制时,club细胞会被委以重任,成为肺泡干细胞再生的主要来源。
那么是何种信号通路调控club细胞及BASCs向AT2细胞发生细胞命运转变?这两种不同干细胞产生AT2细胞的分子调控机制是否一样?为了探明这一点,研究人员首先从损伤4周的Scgb1a1-CreER;Sftpc-DreER;R26-TLR工具小鼠中分选了Club-tracer、BASC-tracer和AT2-tracer标记细胞进行单细胞转录组测序分析(scRNA-seq)。UMAP分析显示Club-tracer标记的细胞可以被划分为7个细胞亚群(cell cluster),这些细胞亚群具有不同的基因表达特点,可依次划为AT2细胞群、Club细胞群、Basal-like细胞群、Cldn18highPreAT1细胞群、Fstl1+transient细胞群、transient细胞群和H2-K1+club细胞群。拟时序分析揭示这些细胞亚群之间具有紧密的谱系联系,club细胞和H2-K1+club细胞作为起点可转变为Basal-like和Fstl1+ transient细胞,也可以通过transient细胞分化为AT2细胞和Cldn18highPreAT1细胞。另一方面,UMAP分析显示根据基因表达BASC-tracer标记的细胞也可被划分为7个细胞亚群,包括Cldn4highPreAT1细胞群、Cxcl15highAT2细胞群、BASC-like 细胞群、Chil1highAT2细胞群、EreghighAT2细胞群、transient细胞群和AT1细胞群。这些细胞亚群在基因表达谱系上具有连贯性,拟时序分析显示细胞分化谱系起始于BASC-like细胞,然后经过transient细胞可依次分化为Cxcl15highAT2、EreghighAT2、Cldn4highPreAT1及AT1细胞,或经Cxcl15highAT2可分化为Chil1highAT2细胞。通过对Club-tracer、BASC-tracer和AT2-tracer中的AT2细胞进行对比分析,发现BASC-tracer中的AT2细胞在基因表达上更接近于AT2-tracer中的AT2细胞。
通过进一步分析,研究人员发现Notch信号通路在club细胞向AT2细胞转分化及BASCs向AT2转分化过程中具有下调趋势,暗示Notch信号通路可能在细胞命运转变中发挥调控作用。为进一步体内验证,研究人员结合双同源重组谱系示踪技术与条件性基因敲除技术构建了Scgb1a1-CreER;Sftpc-DreER;R26-TLR;Rbpjflox/flox工具小鼠(实验组)和Scgb1a1-CreER;Sftpc-DreER;R26-TLR;Rbpjflox/+工具小鼠(对照组),可实现在体内示踪Club-tracer、BASC-tracer和AT2-tracer的同时实现在club细胞和BASCs中特异性敲除Notch信号通路。结合博来霉素损伤模型,研究人员发现与对照组相比,敲除组中club细胞贡献的AT2细胞比例显著性降低,而BASCs贡献的AT2细胞比例显著性上升。此实验证实Notch通路在club及BASCs向AT2转分化过程中发挥重要作用,并且属于两种截然相反的调控作用。为进一步研究Notch通路激活是否会影响BASCs向AT2细胞转分化,研究人员利用双同源重组谱系示踪技术构建了Scgb1a1-CreER;Sftpc-DreER;R26-RL-NICD-GFP遗传工具,可实现对BASCs特异性标记并过表达NICD。结合博来霉素损伤模型,研究人员发现与对照组(Scgb1a1-CreER;Sftpc-DreER;R26-RL-GFP)相比,过表达Notch会显著抑制BASCs向AT2细胞转分化。以上实验证明club细胞可作为AT2细胞的再生起源,并且首次在功能层面上揭示club细胞与BASCs属于两种不同细胞类型,不仅具有不同基因表达特点,而且在向AT2细胞分化过程中受Notch通路相反的调控。
综上,该研究通过构建精准的双同源重组谱系示踪技术系统严格地解析了肺损伤后肺泡上皮干细胞的再生起源,并揭示其分子调控机制,为肺脏干细胞和修复再生领域提供新技术和新方法,也为肺脏疾病的临床治疗研究提供新的治疗靶点和重要研究基础。
国科大杭州高等研究院/中国科学院分子细胞科学卓越创新中心周斌研究组副研究员刘扩、上海科技大学博士生孟鑫凤和中国科学院分子细胞科学卓越创新中心博士生刘子鑫为该论文共同第一作者。周斌研究员为该论文通讯作者。该研究得到分子细胞卓越中心动物实验技术平台和细胞分析技术平台及中国科学院上海营养与健康研究所细胞分析技术平台的大力支持。该工作得到中国科学院、国家自然科学基金委、科技部、上海市科委、国科大杭州高等研究院、新基石科学基金会等支持。
文章链接:https://doi.org/10.1016/j.cell.2024.03.010
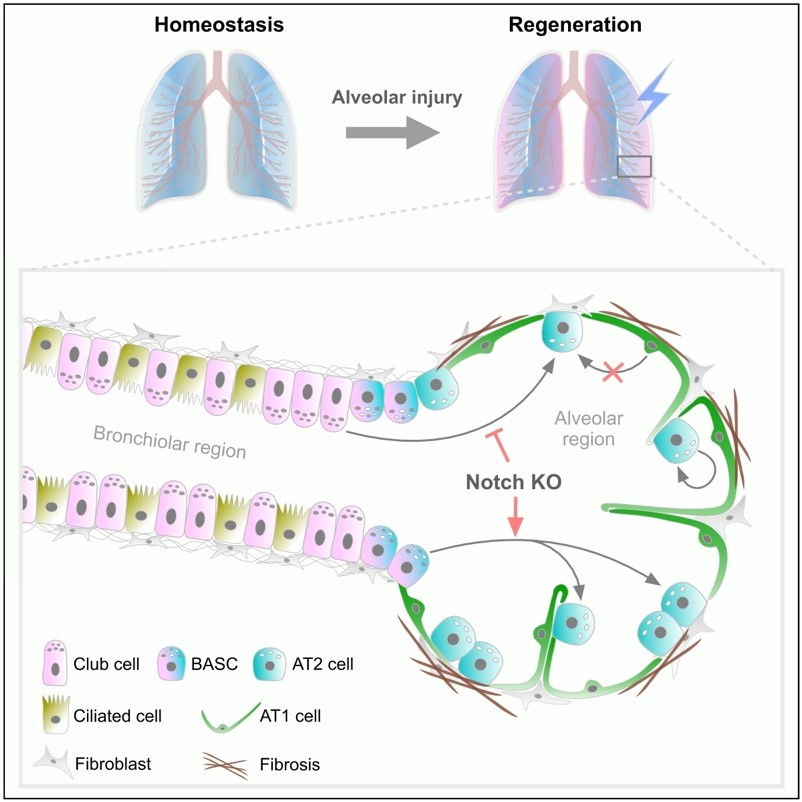
图注:肺脏损伤后AT2细胞的再生起源
利用双同源重组酶介导的遗传谱系示踪技术特异性标记各种不同肺上皮细胞,发现新生AT2细胞除了来源于自我更新外还会起源于club细胞与BASCs,而非AT1细胞。Notch信号通路在club细胞和BASCs向AT2细胞发生命运转变中发挥截然相反的作用,即敲除Notch抑制club细胞向AT2细胞命运转变,但促进BASCs向AT2细胞命运转变。

科普漫画